(MATERIAL CORRESPONDIENTE AL DESARROLLO DE LA MATERIA "NEUROANATOMIA" CORRESPONDIENTE AL PRIMER CURSO DE LA CARRERA DE MEDICINA DE LA UNISAL Y AL CUARTO CURSO DE LA CARRERA DE PSICOLOGIA DE LA UNI)
Generalidades
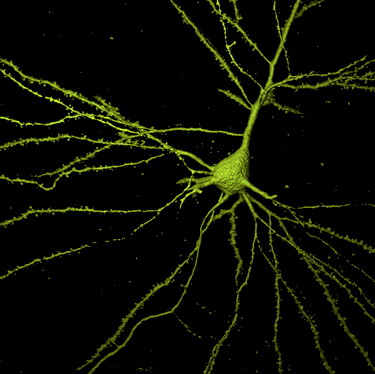
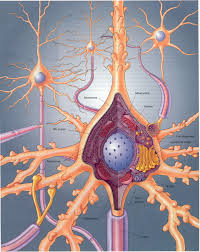
Las neuronas son un tipo de células del sistema nervioso cuya principal característica es la excitabilidad de su membrana plasmática; están especializadas en la recepción de estímulos y conducción del impulso nervioso (en forma de potencial de acción) entre ellas o con otros tipos celulares, como por ejemplo las fibras musculares de la placa motora. Altamente diferenciadas, la mayoría de las neuronas no se dividen una vez alcanzada su madurez; no obstante, una minoría sí lo hace. Las neuronas presentan unas características morfológicas típicas que sustentan sus funciones: un cuerpo celular o «pericarion», central; una o varias prolongaciones cortas que generalmente transmiten impulsos hacia el soma celular, denominadas dendritas; y una prolongación larga, denominada axón o «cilindroeje», que conduce los impulsos desde el soma hacia otra neurona u órgano diana.
La neurogénesis en seres adultos, ha sido descubierta apenas en el último tercio del siglo XX. Hasta hace pocas décadas se creía que, a diferencia de la mayoría de las otras células del organismo, las neuronas normales en el individuo maduro no se regeneraban, excepto las células olfatorias. Los nervios mielinados del sistema nervioso periférico también tienen la posibilidad de regenerarse a través de la utilización del neurolema, una capa formada de los núcleos de las células de Schwann.
Historia
A principios del siglo XX, Santiago Ramón y Cajal situó por vez primera a las neuronas como elementos funcionales del sistema nervioso. Cajal propuso que actuaban como entidades discretas que, comunicándose unas con otras, establecían una especie de red mediante conexiones especializadas o espacios. Esta idea, opuesta a la defendida por Camillo Golgi, que propugnaba la continuidad de la red neuronal (es decir, que negaba que las neuronas fueran entes discretos interconectados), es reconocida como la doctrina de la neurona, uno de los elementos centrales de la neurociencia moderna. A fin de observar al microscopio la histología del sistema nervioso, Cajal empleó tinciones de plata (con sales de plata) de cortes histológicos para microscopía óptica, desarrollados por Golgi y mejorados por el propio Cajal. Dicha técnica permitía un análisis muy preciso, a nivel celular, incluso de un tejido tan denso como es el cerebral.
Morfología
Una neurona típica consta de: un núcleo voluminoso central, situado en el soma; un pericarion que alberga los orgánulos celulares típicos de cualquier célula eucariota; y neuritas (esto es, generalmente un axón y varias dendritas) que emergen del pericarion.
Núcleo
Situado en el cuerpo celular, suele ocupar una posición central y ser muy conspicuo, especialmente en las neuronas pequeñas. Contiene uno o dos nucléolos prominentes, así como una cromatina dispersa, lo que da idea de la relativamente alta actividad transcripcional de este tipo celular. La envolutura nuclear, con multitud de poros nucleares, posee una lámina nuclear muy desarrollada. Entre ambos puede aparecer el cuerpo accesorio de Cajal, una estructura esférica de en torno a 1 ¼m de diámetro que corresponde a una acumulación de proteínas ricas en los aminoácidos arginina y tirosina.
Pericarion
Rico en ribosomas libres y adheridos al retículo endoplasmático rugoso, lo que da lugar a unas estructuras denominadas grumos de Nissl que, al microscopio óptico, se observan como grumos basófilos, y, al electrónico, como apilamientos de cisternas del retículo endoplasmático. Tal abundancia de los orgánulos relacionados en la síntesis proteica se debe a la alta tasa biosintética del pericarion.
El aparato de Golgi es escaso en el pericarion. Existen lisosomas primarios y secundarios (estos últimos, ricos en lipofuscina, pueden marginar al núcleo en individuos de edad avanzada debido a su gran aumento). Las mitocondrias, pequeñas y redondeadas, poseen habitualmente crestas longitudinales.
En cuanto al citoesqueleto, el pericarion es rico en microtúbulos (clásicamente, de hecho, denominados neurotúbulos, si bien son idénticos a los microtúbulos de células no neuronales) y filamentos intermedios (denominados neurofilamentos por la razón antes mencionada).
Dendritas
Las dendritas son ramificaciones que proceden del soma neuronal que consisten en proyecciones citoplasmáticas envueltas por una membrana plasmática sin envuelta de mielina. En ocasiones, poseen un contorno irregular, desarrollando espinas. Sus orgánulos y componentes característicos son: muchos microtúbulos y pocos neurofilamentos, ambos dispuestos en haces paralelos; muchas mitocondrias; grumos de Nissl, más abundantes en la zona adyacente al soma; retículo endoplasmático liso, especialmente en forma de vesículas relacionadas con la sinapsis.
Axón
El axón es una prolongación del soma neuronal recubierta por una o más células de Schwann en el sistema nervioso periférico de vertebrados, con producción o no de mielina. Puede dividirse, de forma centrífuga al pericarion, en: cono axónico, segmento inicial, resto del axón.
* Cono axónico. Adyacente al pericarion, es muy visible en las neuronas de gran tamaño. En él se observa la progresiva desaparición de los grumos de Nissl y la abundancia de microtúbulos y neurfilamentos que, en esta zona, se organizan en haces paralelos que se proyectarán a lo largo del axón.
* Segmento inicial. En él comienza, de existir, la mielinización externa. En el citoplasma, a esa altura se detecta una zona rica en material electronodenso en continuidad con la membrana plasmática, constituido por material filamentoso y partículas densas; se asume que interviene en la generación del potencial de acción que transmitirá la señal sináptica. En cuanto al citoesqueleto, posee esta zona la organización propia del resto del axón. Los microtúbulos, ya polarizados, poseen la proteína Ï pero no la proteína MAP-2.
* Resto del axón. En esta sección comienzan a aparecer los nódulos de Ranvier y las sinapsis.
Función de las neuronas
Las neuronas tienen la capacidad de comunicarse con precisión, rapidez y a larga distancia con otras células, ya sean nerviosas, musculares o glandulares. A través de las neuronas se transmiten señales eléctricas denominadas impulsos nerviosos.
Estos impulsos nerviosos viajan por toda la neurona comenzando por las dendritas, y pasa por toda la neurona hasta llegar a los botones terminales, que pueden conectar con otra neurona, fibras musculares o glándulas. La conexión entre una neurona y otra se denomina sinapsis.
Las neuronas conforman e interconectan los tres componentes del sistema nervioso: sensitivo, integrador o mixto y motor; De esta manera, un estímulo que es captado en alguna región sensorial entrega cierta información que es conducida a través de las neuronas y es analizada por el componente integrador, el cual puede elaborar una respuesta, cuya señal es conducida a través de las neuronas. Dicha respuesta es ejecutada mediante una acción motora, como la contracción muscular o secreción glandular.
El impulso nervioso
Las neuronas transmiten ondas de naturaleza eléctrica originadas como consecuencia de un cambio transitorio de la permeabilidad en la membrana plasmática. Su propagación se debe a la existencia de una diferencia de potencial o potencial de membrana (que surge gracias a las concentraciones distintas de iones a ambos lados de la membrana, según describe el potencial de Nernst) entre la parte interna y externa de la célula (por lo general de -70 mV). La carga de una célula inactiva se mantiene en valores negativos (el interior respecto al exterior) y varía dentro de unos estrechos márgenes. Cuando el potencial de membrana de una célula excitable se despolariza más allá de un cierto umbral ( de 65mV a 55mV app) la célula genera (o dispara) un potencial de acción. Un potencial de acción es un cambio muy rápido en la polaridad de la membrana de negativo a positivo y vuelta a negativo, en un ciclo que dura unos milisegundos.
Neurosecreción
Las células neurosecretoras son neuronas especializadas en la secreción de sustancias que, en vez de ser vertidas en la hendidura sináptica, lo hacen en capilares sanguíneos, por lo que sus productos son transportados por la sangre hacia los tejidos diana; esto es, actúan a través de una vía endocrina. Esta actividad está representada a lo largo de la diversidad zoológica: se encuentra en crustáceos, insectos, equinodermos, vertebrados, etc.
Transmisión de señales entre neuronas
Un sistema nervioso procesa la información siguiendo un circuito más o menos estándar. La señal se inicia cuando una neurona sensorial, generalmente asociada a un órgano de los sentidos, recoge información. Su axón se denomina fibra aferente. Esta neurona sensorial transmite la información a otra aledaña, de modo que acceda un centro de integración del sistema nervioso del animal. Las interneuronas, situadas en dicho sistema, transportan la información a través de sinapsis. Finalmente, si debe existir respuesta, se excitan neuronas eferentes que controlan músculos, glándulas u otras estructuras anatómicas. Las neuronas aferentes y eferentes, junto con las interneuronas, constituyen el circuito neuronal.
La sinapsis es el proceso esencial en la comunicación neuronal y constituye el lenguaje básico del sistema nervioso. Afortunadamente, las semejanzas de los mecanismos sinápticos son mucho más amplias que las diferencias, asociadas éstas a la existencia de distintos neurotransmisores con características particulares.
Elliot en 1904 fue el primero que sugirió la posibilidad de que la información era transferida de una neurona a otra por la liberación de una sustancia química desde las fibras nerviosas; Loewi es, sin embargo, el primero que mostró la existencia de una sustancia química en el líquido perfundido con la estimulación del nervio vago y fue su colaborador Navratil quien más tarde demostró que esta sustancia era la acetilcolina.
La sinapsis es un punto de machimbre o de enlace entre dos neuronas, la presináptica y la postsináptica. Las fibras nerviosas actúan como terminales de bujías eléctricas de los motores de explosión. Hay una luz o una brecha sináptica entre los terminales, brecha sináptica donde descargan vesículas sinápticas que difunden, ayudan a que ocurran reacciones físicas y químicas, recapturan los neurotransmisores ya usados y propagan potencial eléctrico desde una pared o membrana de la brecha o hendidura, la de la neurona presináptica, a la pared o membrana de la otra, la postsináptica.
 En su extremo, el axon de los nervios se ramifica en muchos terminales pequeños que llegan a estar en contacto estrecho con las dendritas de otras neuronas. Al contacto entre dos neuronas se le llama sinapsis. El axon y la dendrita nunca se tocan. Siempre hay un pequeño vacio llamado hendidura sinaptica. Cuando la señal electrica llega a un terminal nervioso, hace que el nervio libere neurotransmisores. Los neurotransmisores son agentes quimicos que viajan una corta distancia hasta las dendritas mas próximas.
En su extremo, el axon de los nervios se ramifica en muchos terminales pequeños que llegan a estar en contacto estrecho con las dendritas de otras neuronas. Al contacto entre dos neuronas se le llama sinapsis. El axon y la dendrita nunca se tocan. Siempre hay un pequeño vacio llamado hendidura sinaptica. Cuando la señal electrica llega a un terminal nervioso, hace que el nervio libere neurotransmisores. Los neurotransmisores son agentes quimicos que viajan una corta distancia hasta las dendritas mas próximas.
A la neurona que libera el neurotransmisor se le llama neurona presinaptica. A la neurona receptora de la señal se le llama neurona postsinaptica. Dependiendo del tipo de neurotransmisor liberado, las neuronas postsinapticas son estimuladas (excitadas) o desestimuladas (inhibidas). Cada neurona se comunica con muchas otras al mismo tiempo. Puesto que una neurona puede enviar o no un estimulo, su comportamiento siempre se basa en el equilibrio de influencias que la excitan o la inhiben en un momento dado. Las neuronas son capaces de enviar estimulos varias veces por segundo.
Existen dos tipos de sinapsis: las electricas y las quimicas.
En las sinapsis electricas los procesos pre y postsináptico son continuos (2 nm entre ellos) debido a la unión citoplasmática por moléculas de proteínas tubulares a través de las cuales transita libremente el agua, pequeños iones y moléculas por esto el estímulo es capaz de pasar directamente de una célula a la siguiente sin necesidad de mediación química (Barr, 1994). Corresponden a uniones de comunicación entre las membranas plasmáticas de los terminales presináptico y postsinápticos, las que al adoptar la configuración abierta permiten el libre flujo de iones desde el citoplasma del terminal presinático hacia el citoplasma del terminal postsináptico.
La sinapsis eléctrica ofrece una vía de baja resistencia entre neuronas, y hay un retraso mínimo en la transmisión sináptica porque no existe un mediador químico. En este tipo de sinapsis no hay despolarización y la dirección de la transmisión está determinada por la fluctuación de los potenciales de membrana de las células interconectadas (Bradford, 1988).
La mayoría de las sinapsis son de tipo químico, en las cuales una sustancia, el neurotransmisor hace de puente entre las dos neuronas, se difunde a través del estrecho espacio y se adhiere a los receptores, que son moléculas especiales de proteínas que se encuentran en la membrana postsináptica (Bradford, 1988).
La energía requerida para la liberación de un neurotransmisor se genera en la mitocondria del terminal presináptico. La unión de neurotransmisores a receptores de la membrana postsinápticas produce cambios en la permeabilidad de la membrana. La naturaleza del neurotransmisor y la molécula del receptor determina si el efecto producido será de excitación o inhibición de la neurona postsináptica (Barr, 1994). Se han descrito varias formas de sinapsis según las estructuras implicadas. (Bradford, 1988).
 Axosomática: Sinapsis entre un axón y un soma.
Axosomática: Sinapsis entre un axón y un soma.
Axodendrítica: Sinapsis ocurrida entre un axón y una dendrita.
Axoespinodendrítica: Sinapsis entre un axón y una espina dendrítica.
Axoaxónica: Sinapsis entre dos axones.
Dendrodendrítica: Sinapsis ocurrida entre dos dendritas.
Somatosomática: Sinapsis entre dos somas.
Dendrosomática: Sinapsis entre un soma y una dendrita.
Clasificación
Aunque el tamaño del cuerpo celular puede ser desde 5 hasta 135 micrómetros, las prolongaciones o dendritas pueden extenderse a una distancia de más de un metro. El número, la longitud y la forma de ramificación de las dendritas brindan un método morfológico para la clasificación de las neuronas.
Según la forma y el tamaño
Según el tamaño de las prolongaciones, los nervios se clasifican en:
* Poliédricas: como las motoneuronas del asta anterior de la médula.
* Fusiformes: como las células de doble ramillete de la corteza cerebral.
* Estrelladas: como las neuronas aracniformes y estrelladas de la corteza cerebral y las estrelladas, en cesta y Golgi del cerebelo.
* Esféricas: en ganglios raquídeos, simpáticos y parasimpáticos
* Piramidales: presentes en la corteza cerebral.
Según la polaridad
Según el número y anatomía de sus prolongaciones, las neuronas se clasifican en:
* Neuronas monopolares o unipolares: son aquéllas desde las que nace sólo una prolongación que se bifurca y se comporta funcionalmente como un axón salvo en sus extremos ramificados en que la rama periférica reciben señales y funcionan como dendritas y transmiten el impulso sin que este pase por el soma neuronal. Son típicas de los ganglios de invertebrados y de la retina.
* Neuronas bipolares: poseen un cuerpo celular alargado y de un extremo parte una dendrita y del otro el axón (solo puede haber uno por neurona). El núcleo de este tipo de neurona se encuentra ubicado en el centro de ésta, por lo que puede enviar señales hacia ambos polos de la misma. Ejemplos de estas neuronas se hallan en las células bipolares de la retina (conos y bastones), del ganglio coclear y vestibular, estos ganglios son especializados de la recepción de las ondas auditivas y del equilibrio.
* Neuronas multipolares: tienen una gran cantidad de dendritas que nacen del cuerpo celular. Ese tipo de células son la clásica neurona con prolongaciones pequeñas (dendritas) y una prolongación larga o axón. Representan la mayoría de las neuronas. Dentro de las multipolares, distinguimos entre las que son de tipo Golgi I, de axón largo, y las de tipo Golgi II, que no tienen axón o éste es muy corto. Las neuronas de proyección son del primer tipo, y las neuronas locales o interneuronas del segundo.
* Neuronas pseudounipolares: son aquéllas en las cuales el cuerpo celular tiene una sola dendrita o neurita, que se divide a corta distancia del cuerpo celular en dos ramas, motivo por cual también se les denomina pseudounipolares (pseudos en griego significa “falso”), una que se dirige hacia una estructura periférica y otra que ingresa en el sistema nervioso central. Se hallan ejemplos de esta forma de neurona en el ganglio de la raíz posterior.
* Neuronas anaxónicas: son pequeñas. No se distinguen las dendritas de los axones. Se encuentran en cerebro y órganos especiales de los sentidos.
Según las características de las neuritas
De acuerdo a la naturaleza del axón y de las dendritas, clasificamos a las neuronas en:
* Axón muy largo o Golgi de tipo I. El axón se ramifica lejos del pericarion. Con axones de hasta 1 m.
* Axón corto o Golgi de tipo II. El axón se ramifica junto al soma celular.
* Sin axón definido. Como las células amacrinas de la retina.
* Isodendríticas. Con dendritas rectilíneas que se ramifican de modo que las ramas hijas son mas laÅ?gas que las madres.
* Idiodendríticas. Con las dendritas organizadas dependiendo del tipo neuronal; por ejemplo, como las células de Purkinje del cerebelo.
* Alodendríticas. Intermedias entre los dos tipos anteriores.
Según el mediador químico
Las neuronas pueden clasificarse, según el mediador químico, en:
* Colinérgicas. Liberan acetilcolina.
* Noradrenérigicas. Liberan norepinefrina.
* Dopaminérgicas. Liberan dopamina.
* Serotoninérgicas. Liberan serotonina.
* GABAérgicas. Liberan GABA, es decir, ácido γ-aminobutírico.
Doctrina de la neurona
La doctrina de la neurona, establecida por Santiago Ramón y Cajal a finales del siglo XIX, es el modelo aceptado hoy en neurofisiología. Consiste en aceptar que la base de la función neurológica radica en las neuronas como entidades discretas, cuya interacción, mediada por sinapsis, conduce a la aparición de respuestas complejas. Cajal no solo postuló este principio, sino que lo extendió hacia una «ley de la polarización dinámica», que propugna la transmisión unidireccional de información (esto es, en un sólo sentido, de las dendritas hacia los axones). No obstane, esta ley no siempre se cumple. Por ejemplo, las células gliales pueden intervenir en el procesamiento de información, e, incluso, las efapsis o sinapsis eléctricas, mucho más abundantes de lo que se creía, presentan una transmisión de información directa de citoplasma a citoplasma. Más aún: las dendritas pueden dirigir una señal sináptica de forma centrífuga al soma neuronal, lo que representa una transmisión en el sentido opuesto al postulado, de modo que sean los axones los que reciban de información (aferencia).
Redes neuronales
Una red neuronal se define como una población de neuronas físicamente interconectadas o un grupo de neuronas aisladas que reciben señales que procesan a la manera de un circuito reconocible. La comunicación entre neuronas, que implica un proceso electroquímico, implica que, una vez que una neurona es excitada a partir de cierto umbral, ésta se despolariza transmitiendo a través de su axón una señal que excita a neuronas aledañas, y así sucesivamente. El sustento de las capacidad del sistema nervioso, por tanto, radica en dichas conexiones. En oposición a la red neuronal, se habla de circuito neuronal cuando se hace mención a neuronas que se controlan dando lugar a una retroalimentación («feedback»), como define la cibernética.
Cerebro y neuronas
El número de neuronas en el cerebro varía drásticamente según la especie estudiada. Se estima que cada cerebro humano posee en torno a 1011 neuronas: es decir, unos cien mil millones. No obstante, Caenorhabditis elegans, un gusano nematodo muy empleado como animal modelo, posee sólo 302.; y la mosca de la fruta, Drosophila melanogaster, unas 300.000, que bastan para permiterle exhibir conductas complejas. La fácil manipulación en el laboratorio de estas especies, cuyo ciclo de vida es muy corto y cuyas condiciones de cultivo porco exigentes, permiten a los investigadores científicos emplearlas para dilucidar el funcionamiento neuronal, puesto que el mecanismo básico de la actividad neuronal es común al de nuestra especie.
Redes neuronales artificiales
El conocimiento de las redes neuronales biológicas ha dado lugar a un diseño empleado en inteligencia artificial. Estas redes funcionan porque cada neurona recibe una serie de entradas a través de interconexiones y emite una salida. Esta salida viene dada por tres funciones: una función de propagación que por lo general consiste en el sumatorio de cada entrada multiplicada por el peso de su interconexión; una función de activación, que modifica a la anterior y que puede no existir, siendo en este caso la salida la misma función de propagación; y una función de transferencia, que se aplica al valor devuelto por la función de activación. Se utiliza para acotar la salida de la neurona y generalmente viene dada por la interpretación que queramos darle a dichas salidas.